Glutamato.
Aunque existen diferentes clases de neuronas, las más abundantes en el sistema nervioso central son las neuronas glutamatérgicas y las GABAérgicas. Las primeras producen despolarización de la membrana postsináptica, que es la característica de la sinapsis excitatorias; mientras que las segundas producen la hiperpolarización rasgo particular de las sinapsis inhibitorias; Se estima que las sinapsis glutamatérgicas que usan el GLU para la neurotransmisión constituyen 70% del total y usualmente están situadas sobre las espinas dendríticas, estructuras que aumentan la superficie de contacto entre células.
El GLU es almacenado en vesículas (100 mM) y es liberado en la sinapsis por exocitosis Ca+2 dependiente en la membrana post-sináptica el GLU reconoce al menos dos tipos de receptores, los metabotrópicos de GLU y los receptores ionotrópicos de GLU; los mGluR son una familia de receptores asociados con proteínas G y median mecanismos de señalización activando diferentes cascadas de señalización. Se conocen 8 tipos de receptores metabotrópicos de GLU mGluR1-8, organizados en tres grupos (grupo I, II y III) teniendo en cuenta la homología de sus secuencias, la respuesta farmacológica a los agonistas y el acoplamiento a sistemas intracelulares de transducción de las señales.
Los iGluR han sido clasificados en tres poblaciones diferentes, cada una definida por la activación selectiva con diferentes análogos estructurales del GLU: NMDA (N-metil-D-Aspartato), AMPA (α-amino-3-hidroxil-5-metil-4-isoxazol-propionato) y KAIN (ácido Kainico); los iGluR están asociados a canales catiónicos selectivos que permiten la entrada primariamente de Ca2+ y Na+ a la célula, mientras que el K+ efluye de ella a través del mismo canal.
🧠Metabolismo del Glutamato.
El GLU como neurotransmisor proviene metabólicamente de los esqueletos de carbono de la glucosa que atraviesan la barrera hematoencefálica y del ciclo glutamato/glutamina lo que crea una estrecha interacción entre neurona y astrocitos los sustratos que se sintetizan y se transportan favoreciendo a la actividad sináptica. En la actividad glutamatérgica el glutamato es liberado e interactúa con diferentes receptores en la membrana post sináptica y su exceso es tomado por los astrocitos en un proceso sodio dependiente y convertido en GLN o en α–cetoglutarato.La GLN es luego liberada por los astrocitos y recapturada por las neuronas en donde actúa la enzima PAG de-aminándola y transformándola en GLU, que puede ser de nuevo usado en la neurotransmisión, o en el metabolismo oxidativo, de forma similar a como lo hacen las células hepáticas.
Giménez, C; Zafra, F; Aragón, C. (2018). Pathophysiology of the glutamate and the glycine transporters: new therapeutic targets. Journal Neurology. 67 (12), 491-504.https://doi.org/10.33588/rn.6712.2018067

La glutamina se importa a la neurona glutamatérgica (A) y se convierte en glutamato por acción de la glutaminasa. El glutamato se concentra entonces en las vesículas por el transportador de glutamato vesicular. Al liberarse hacia la sinapsis el glutamato puede interactuar con los conductos de receptores ionotrópicos AMPA y NMDA (AMPAR, NMDAR) en la densidad postsináptica (PSD) y con receptores metabotrópicos (MGluR) en la célula postsináptica (B). La transmisión sináptica termina por transporte activo del glutamato hacia una célula de la glía vecina (C) por un transportador de glutamato. Se sintetiza glutamina a partir de éste por acción de la sintetasa de glutamina y se exporta al axón glutamatérgico. En (D) se muestra un modelo del conducto del complejo receptor de NMDA constituido por una proteína tetramérica que se torna permeable a Na+ y Ca2+ cuando se une a una molécula de glutamato.
Katzung, B; Masters, S; & Trevor, A. (2012). Esquema de una sinapsis de glutamato. [Esquema]. Recuperado del libro Farmacología Básica y Clínica 12a Edición.
🧠Aminoácidos Excitadores-Conclusiones.
Ácido 𝛄-Aminobutírico.

Es un neurotransmisor ampliamente distribuido en las neuronas del córtex cerebral.
Su concentración varía de 2 a 5 mcmol/g en la sustancia gris, siendo la máxima (10 mcmol/g) en la región nigroestriada (esta vía es una de las rutas cerebrales de transmisión dopaminérgica).
Es un tipo de sustancia que es utilizada por las neuronas del sistema nervioso a la hora de comunicarse entre sí a través de unos espacios (llamados espacios sinápticos) por los cuales se conectan entre ellas.
Realiza algunas funciones que otros neurotransmisores no hacen. Su función es la de ser un neurotransmisor inhibitorio.
Se encarga de inhibir o reducir la actividad neuronal jugando un papel importante en el comportamiento, la cognición y la respuesta del cuerpo frente al estrés.
Las investigaciones sugieren que el GABA ayuda a controlar el miedo y la ansiedad cuando las neuronas se sobreexcitan.
Por otro lado, los niveles bajos de este neurotransmisor se asocian a trastornos de ansiedad, problemas para dormir, depresión y esquizofrenia. También se ha constatado que las neuronas jóvenes son más excitables que las antiguas, esto es debido a la función que ejerce el GABA sobre las últimas.
El GABA Contribuye al control motor, la visión, regula la ansiedad, entre otras funciones corticales. Existen distintos fármacos que aumentan los niveles de GABA en el cerebro y se utilizan para tratar la epilepsia, la enfermedad de Huntington o para calmar la ansiedad (por ejemplo, las benzodiazepinas). El GABA es un neurotransmisor, participa en la comunicación entre neuronas.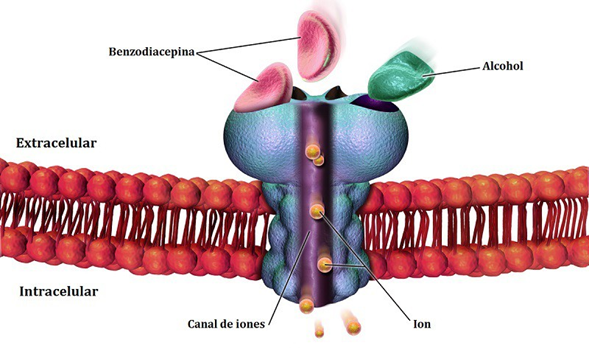
🧠Síntesis.
El GABA se sintetiza a partir de la descarboxilación del glutamato gracias a la acción de la enzima glutamato descarboxilasa (GAD), un proceso que ocurre en las neuronas gabaérgicas en el cerebelo, los ganglios basales y muchas áreas de la corteza cerebral, también en la médula espinal. (Si se inhibe la síntesis de este neurotransmisor se producen ataques convulsivos).
🧠Receptores.
Se han identificado dos tipos de receptores para el ácido y- aminobutírico: GABAA y GABAB. Un tercer tipo de receptor (GABAC) se considera hoy en día un subtipo de receptor GABAA.
🧠GABAA

Los receptores GABAA pertenecen a la misma familia que los receptores para otros transmisores como la glicina, ácido nicotínico y serotonina.
Son pentámeros. Se han clonado varias subunidades, tiene una estructura «α-β-α-β-γ» que conforma un poro central (canal de aniones Cl-), al localizarse en las membranas de las neuronas post-sinápticas, su activación tras la unión de su ligando fisiológico aumenta la permeabilidad de los aniones Cl-, hiperpolarizandose la membrana, con la consiguiente disminución de la excitabilidad.
Los receptores del GABAA se localizan tanto en sinapsis como en ubicaciones extrasinápticas. Estas últimas dan lugar a una inhibición «a distancia», una neuromodulación alejada del punto donde se segregó la molécula. Los receptores extrasinápticos del GABAA tienen una estructura cuaternaria «α4/α5/α6/δ». Estos receptores extrasinápticos son muy sensibles a los anestésicos y al etanol. Además tienen una mayor afinidad por el GABAA y muestran menos desensibilización. Gaboxadol (THIP), un agonista parcial del receptor GABAA, muestra afinidad por la subunidad δ.
Ligandos de receptores GABAA :
-Agonistas moduladores alostéricos positivos: etanol, barbitúricos, benzodiazepinas, el hidrato clorar, la valeriana común y los anestésicos inhalados volátiles.
-Antagonistas o moduladores alostéricos negativos: furosemida, flumazenil, gabazina, enantotoxina, bicucolina, picotoxina.
🧠GABAB

Los receptores GABAB se localizan pre- y post-sinápticamente.
Los receptores GABAB pertenecen a la clase de proteínas C acopladas con proteínas G que regulan la permeabilidad transmembrana a los cationes Ca 2+. Consecuencia: disminuye la excitabilidad post-sináptica, con subsiguiente inhibición de la actividad enzimática adenilato-ciclasa.
El receptor GABAB está formado por dos dímeros: GABAB1 y GABAB2. Las colas de estos dos dímeros atraviesan la bicapa lipídica de la membrana. Cuando el GABA (ligando fisiológico) se engarza con el dominio extracelular de la subunidad GABAB1 se produce un cambio conformacional en la subunidad GABAB2 que activa una vía de señalización celular (vía proteínas G).
Ligandos de receptores GABAB:
Agonistas: bacoflen, propofol, fenibut.
Antagonistas: facofeln y sacolfen.
Inhibidores de recaptura de GABA: hiperforina y tiagabina.
Inhibidores de la GABA transaminasa: gabaculina, fenelzina, ácido valproico, vigabatrina.
Análogos de GABA: pregabalina y gabapentina.
Otros: GABA por sí mismo, L-glutamina, picamilon y progabide.
Glicina
El receptor de glicina (v. Lynch, 2009) se asemeja al receptor GABAA por el hecho de ser un canal de cloruro controlado por ligando, pentamérico y de asa Cys. No existen receptores metabotrópicos específicos de glicina. Se han clonado cinco subunidades del receptor de glicina (a1-4, b) y parece que en el encéfalo adulto la principal forma del receptor está constituida por subunidades a1 y b, aunque que la estequiometría precise de tales subunidades es objeto de debate. La situación en lo que respecta a la glicina es, pues, mucho más sencilla que la del GABA. Se han identificado mutaciones del receptor en ciertos trastornos neurológicos hereditarios asociados a espasmo muscular e hiperexcitabilidad refleja. No hay fármacos de acción terapéutica que actúen específicamente modificando los receptores de glicina, aunque parece evidente que muchos de los compuestos que potencian la activación de los receptores GABAA (como las benzodiacepinas y los fármacos anestésicos) actúan de manera similar sobre los receptores de glicina.
La toxina del tétanos, una toxina bacteriana semejante a la toxina botulínica, actúa de forma selectiva inhibiendo la liberación de glicina por parte de las interneuronas inhibidoras de la médula espinal, dando lugar a una excesiva hiperexcitabilidad refleja y a violentos espasmos musculares (trismo).
La glicina es retirada del espacio extracelular por dos transportadores, GlyT1 y GlyT2, (Eulenburg et al., 2005). El GlyT1 se localiza principalmente en los astrocitos y se expresa en la mayoría de las regiones del SNC. Por su parte, el GlyT2 se expresa en las neuronas glicinérgicas de la médula espinal, el tronco del encéfalo y el cerebelo. Como se ha indicado anteriormente, además de su función como transmisor inhibidor, la glicina actúa también como coagonista con el glutamato en los receptores de NMDA. La inhibición de la captación de glicina a través del GlyT1 da lugar a una elevación de los niveles de glicina extracelular en el encéfalo y, debido a la consiguiente potenciación de las respuestas mediadas por los receptores de NMDA, podría resultar beneficiosa en el tratamiento de la esquizofrenia. Otra posible aplicación de los inhibidores de los transportadores de glicina es su uso como analgésicos.
🧠Aminoácidos Inhibidores-Conclusiones.
- El GABA es el principal transmisor inhibidor del encéfalo.
- Se encuentra distribuido de manera bastante uniforme por todo el encéfalo, pero apenas existe en los tejidos periféricos.
- El GABA se forma a partir del glutamato, mediante la acción de la descarboxilasa del ácido glutámico. El principal mecanismo utilizado para interrumpir su acción es la recaptación, aunque también puede degradarse por desaminación, una reacción catalizada por la GABA transaminasa.
- Existen dos tipos de receptores de GABA: GABAA y GABAB.
- Los receptores GABAA, localizados sobre todo en posición postsináptica, están acoplados directamente a los canales de cloruro, cuya apertura reduce la excitabilidad de la membrana.
- El muscimol es un agonista específico del GABA y el convulsivo bicuculina es un antagonista.
- Otros fármacos que actúan sobre los receptores GABAA y los canales son:
– Las benzodiacepinas, que actúan en un sitio de unión accesorio para facilitar la acción del GABA.
– Convulsivos tales como la picrotoxina, que bloquean el canal aniónico.
– Neuroesteroides, entre ellos los metabolitos endógenos de la progesterona, y otros depresores del SNC, como los barbitúricos y algunos anestésicos generales, que facilitan la acción del GABA. - Los receptores GABAB son receptores heterodiméricos acoplados a proteínas G. Producen inhibición pre- y postsináptica al impedir la apertura de canales de calcio y aumentar la conductancia del K+. El baclofeno es un agonista del receptor GABAB que se emplea para tratar la espasticidad. Por el momento, los antagonistas del GABAB no tienen uso clínico.
- La glicina es un transmisor inhibidor que actúa principalmente en la médula espinal a través de un receptor propio, aunque estructural y funcionalmente es similar al receptor GABAA.
- La estricnina, una sustancia convulsiva, es un antagonista competitivo de la glicina. La toxina tetánica actúa fundamentalmente inhibiendo la liberación de glicina.
Comentarios
Publicar un comentario